


Las neuronas generan y propagan potenciales de acción a lo largo de sus axones, y a continuación transmiten la señal correspondiente a través de una sinapsis mediante el neurotransmisor liberado, que desencadena una reacción en otra neurona o en una célula efectora (p. ej., células musculares, la mayoría de las células exocrinas y endocrinas). Los neurotransmisores permiten que las neuronas se comuniquen entre sí. Los neurotransmisores que se liberan se unen a los receptores de otra neurona. Las neuronas que liberan neurotransmisores se llaman neuronas presinápticas. Las neuronas que reciben señales de neurotransmisores se llaman neuronas postsinápticas. La señal puede estimular o inhibir a la célula receptora, dependiendo del neurotransmisor y el receptor involucrados. Otros factores, como fármacos y trastornos, afectan la comunicación entre las neuronas mediante la modulación de la producción y las acciones de los neurotransmisores, que incluyen
Su liberación, recaptación y degradación
El número y la función de los receptores de neurotransmisores postsinápticos
A veces, las señales entre las neuronas se producen en la dirección inversa (llamada neurotransmisión retrógrada). En tales casos, las dendritas (ramas receptoras de una neurona) en las neuronas postsinápticas liberan neurotransmisores que afectan a los receptores en las neuronas presinápticas. La transmisión retrógrada puede inhibir la liberación de neurotransmisores adicionales en las neuronas presinápticas y ayudar a controlar el nivel de actividad y la comunicación entre las neuronas.
En el sistema nervioso central, las interconexiones son complejas. Un impulso de una neurona a otra puede pasar del
Axón al cuerpo celular
Axón a la dendrita
Cuerpo celular al cuerpo celular
Dendrita a dendrita
Una neurona puede recibir simultáneamente muchos impulsos (excitadores e inhibidores) desde otras neuronas e integrar impulsos simultáneos en distintos patrones de descarga.
Propagación
La propagación del potencial de acción a lo largo del axón es eléctrica, causada por los intercambios de iones de sodio y potasio a través de la membrana axónica. Una neurona particular genera el mismo potencial de acción después de cada estimulo y lo transmite a una velocidad fija a lo largo del axón. La velocidad depende del diámetro del axón y el grado de mielinización y varía de 1 a 4 m/s en las fibras amielínicas pequeñas hasta 75 m/s en las grandes fibras mielínicas. La velocidad de propagación es más alta en las fibras mielínicas porque la cubierta de mielina tiene brechas regulares (nódulos de Ranvier) donde el axón queda expuesto. El impulso eléctrico pasa de un nódulo al siguiente, saltándose el segmento mielínico del axón. Por lo tanto, los trastornos que alteran la cubierta de mielina (p. ej., esclerosis múltiple, síndrome de Guillain-Barré) interfieren con la propagación de los impulsos y causan diversos síntomas neurológicos.
Transmisión
La transmisión de los impulsos es química, causada por la liberación de neurotransmisores específicos desde la terminación nerviosa. Los neurotransmisores difunden a través de la hendidura sináptica y se unen brevemente a receptores específicos en la neurona adyacente o la célula efectora. En función del receptor, la respuesta puede ser excitadora o inhibidora. Por lo general, las neuronas no se tocan entre sí; en cambio, se comunican a través de la transmisión de neurotransmisores mediante las sinapsis. En algunas condiciones, las neuronas cercanas entre sí pueden comunicarse mediante impulsos eléctricos a través de una unión estrecha.
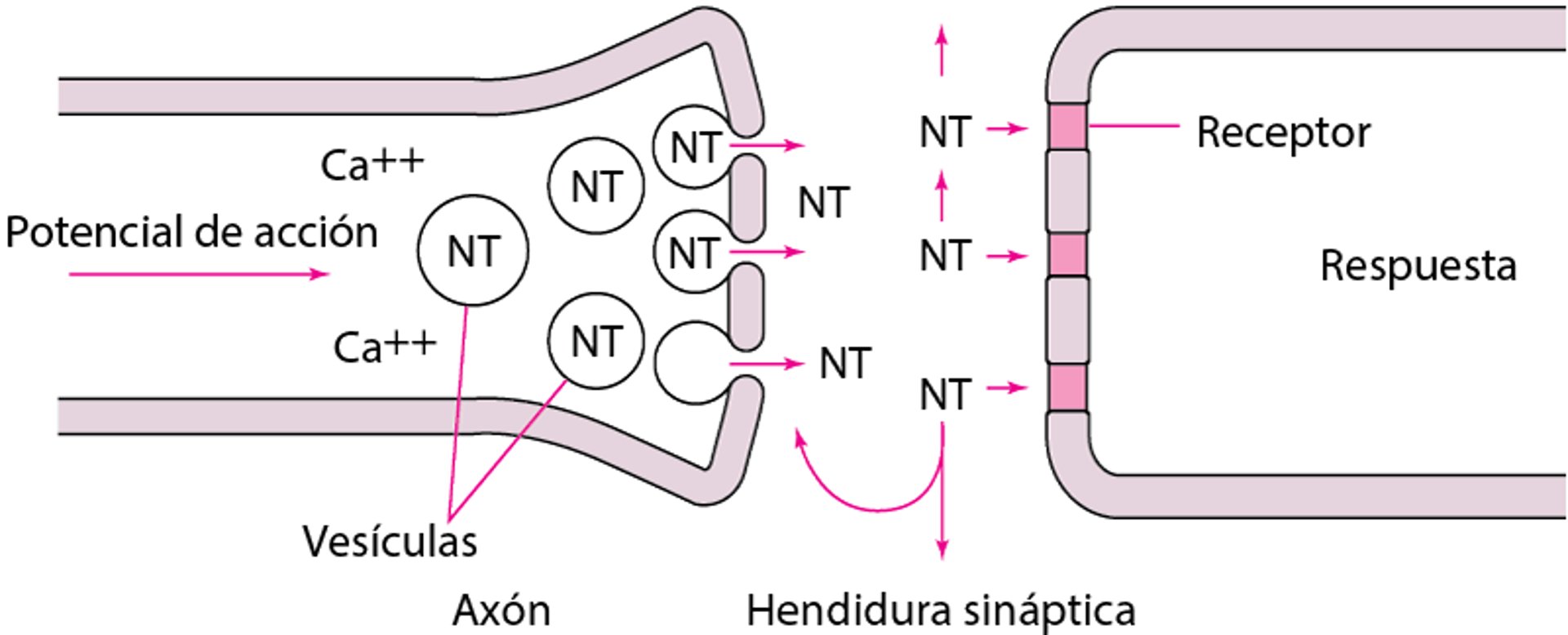
El soma neuronal produce enzimas encargadas de sintetizar a la mayoría de los neurotransmisores, que son almacenados en vesículas en las terminaciones nerviosas (véase figura Neurotransmisión). La cantidad contenida en una vesícula (habitualmente, varios miles de moléculas) es un cuanto. Un potencial de acción de membrana que llega a la terminación abre los canales de calcio del axón; el influjo de calcio libera moléculas de neurotransmisor desde muchas vesículas al fusionar las membranas de la vesícula con la membrana de la terminación nerviosa. La fusión de la membrana genera una apertura a través de la cual las moléculas son expulsadas en la hendidura sináptica a través de la exocitosis.
Hay un tipo de sinapsis, la eléctrica, en la que no participan neurotransmisores; los canales iónicos conectan directamente el citoplasma de las neuronas presinápticas y postsinápticas. Este tipo de transmisión es la más rápida.
Señales excitadoras e inhibidoras
La reacción desencadenada por la liberación de neurotransmisores puede excitar o activar la neurona postsináptica o inhibir o bloquear su actividad. Las neuronas postsinápticas reciben múltiples señales de neurotransmisores y señales eléctricas de muchas neuronas. La neurona receptora finalmente suma los estímulos aferentes y, si se reciben más señales excitadoras, la neurona se activa y envía señales a otras neuronas. Si la suma de las señales es inhibidora, la neurona no se activa y no influye en la actividad de otras neuronas. Esta suma de respuestas se llama sumatoria. Los neurotransmisores facilitan así la comunicación rápida entre las neuronas al modificar la descaga de potenciales de acción.
Otras formas de sumación incluyen
Sumatoria espacial: cuando se reciben múltiples impulsos en diferentes ubicaciones de la neurona y la neurona los suma
Sumatoria temporal: cuando los impulsos se reciben en un corto período de tiempo y luego se suman
Para que una neurona genere una señal y se dispare, debe alcanzar un potencial umbral. Un potencial umbral se produce por un aumento neto en la entrada de sodio en la célula durante el intercambio de iones de sodio y potasio. Cuando suficiente sodio ingresa a la célula, se alcanza el umbral; cuando se alcanza el umbral, se dispara un potencial de acción; este viaja a lo largo de la membrana de la neurona. El umbral debe ser alcanzado para que se genere un potencial de acción.
Neurotransmisión
Los potenciales de acción abren los canales axónicos de calcio (Ca) (no representados). El Ca++ activa la liberación de neurotransmisores (NT) desde las vesículas donde están almacenados. Las moléculas del NT llenan la hendidura sináptica. Algunas se unen a los receptores postsinápticos e inician una respuesta. Las otras son bombeadas de nuevo hacia el axón para su almacenamiento o su difusión hacia los tejidos vecinos. |

Un rasgo característico es la independencia entre la cantidad de neurotransmisor presente en la terminación y la actividad nerviosa, y su contenido se mantiene relativamente constante aunque se modifique la captación de los precursores de estas sustancias o el funcionamiento de las enzimas que intervienen en su síntesis o su destrucción. La estimulación de los receptores presinápticos puede disminuir la síntesis de neurotransmisor, y su bloqueo, incrementarla.
La interacción del neurotransmisor con el receptor debe interrumpirse pronto para finalizar la acción continua del neurotransmisor y/o para permitir la activación rápida y repetida de los receptores. Los neurotransmisores que han interactuado con los receptores pueden sufrir algunas de las situaciones siguientes:
Pueden ser bombeados a gran velocidad de nuevo a las terminaciones nerviosas mediante procesos activos dependientes de ATP (recaptación) para su reciclado o su destrucción.
Pueden ser eliminados por enzimas cerca de los receptores.
Pueden difundir hacia la zona adyacente y ser eliminados.
Los neurotransmisores captados por las terminaciones nerviosas son reempaquetados en gránulos o vesículas en el axón terminal para proceder a su reutilización.
El funcionamiento anormal de estos procesos puede producir una enfermedad clínica. Por ejemplo, se postula que la pérdida de memoria en la enfermedad de Alzheimer implica la insuficiencia del neurotransmisor acetilcolina en las sinapsis, que media la creación de nuevos recuerdos. Ciertos medicamentos (p. ej., donepezilo, galantamina, rivastigmina) bloquean la enzima acetilcolinesterasa (que degrada la acetilcolina) y, por lo tanto, aumentan la cantidad de acetilcolina en la sinapsis. Como resultado, la función de la memoria puede mejorar.
Algunos tipos de neuronas individuales pueden liberar dos o más neurotransmisores diferentes (llamada cotransmisión), por ejemplo, acetilcolina y glutamato. Los neurotransmisores múltiples pueden actuar en una sola neurona postsináptica o afectar múltiples neuronas postsinápticas. La cotransmisión permite una comunicación intrincada entre las neuronas para controlar diferentes eventos en el sistema nervioso central y el sistema nervioso periférico (SNP).
Los neurotransmisores también pueden facilitar más cambios a largo plazo que implican vías adicionales, como cambios en la actividad de genes y proteínas.
Receptores
Los receptores para los neurotransmisores consisten en complejos proteicos que atraviesan la membrana celular. Su naturaleza determina si una sustancia concreta es excitadora o inhibidora. Los receptores que son estimulados continuamente por neurotransmisores o fármacos quedan desensibilizados (regulados en menos); si no son activados por su neurotransmisor o están bloqueados de forma crónica por fármacos, se vuelven hipersensibles (regulados en más). La regulación en menos o en más de los receptores influye fuertemente en el desarrollo de tolerancia y dependencia física. Estos conceptos son particularmente importantes en el trasplante de órganos o tejidos, en el que la denervación priva a los receptores de su neurotransmisor; como resultado, los órganos trasplantados pueden volverse demasiado sensibles a la estimulación nerviosa. Los síntomas de abstinencia pueden explicarse al menos en parte por un fenómeno de rebote debido a la modificación de la afinidad de los receptores o de su densidad.
La mayoría de los neurotransmisores interactúan básicamente con los receptores postsinápticos, pero algunos receptores se localizan en las neuronas presinápticas, lo que proporciona un control delicado sobre la liberación del neurotransmisor.
Una familia de receptores, denominados receptores ionotrópicos (p. ej., N-metil-d-glutamato, cinasa-quiscualato, colinérgicos nicotínicos, de glicina y de ácido gamma-aminobutírico [GABA]), consiste en canales iónicos que se abren cuando se les une el neurotransmisor, lo que facilita una repuesta muy rápida. En la otra familia, denominada receptores metabotrópicos (p. ej., receptores serotoninérgicos, alfa- y beta-adrenérgicos y dopaminérgicos), los neurotransmisores interactúan con proteínas G y activan a otra molécula (segundo mensajero como el cAMP) que cataliza una cadena de acontecimientos cuyo resultado es la fosforilación proteica del calcio o su movilización; las respuestas mediadas por un sistema de segundos mensajeros son más lentas y permiten una regulación más delicada de la respuesta rápida de los neurotransmisores ionotrópicos. Es mucho mayor el número de neurotransmisores que activan receptores específicos que segundos mensajeros.
Neurotransmisores y receptores principales
Un mínimo de 100 sustancias puede actuar como neurotransmisores; unas 18 tienen una gran importancia. Varias adoptan formas un poco diferentes. Los neurotransmisores se pueden agrupar en diferentes clases, como
Moléculas pequeñas (p. ej., glutamato, ácido gamma-aminobutírico, glicina, adenosina, acetilcolina, serotonina, histamina, noradrenalina)
Neuropéptidos (p. ej., endorfinas)
Moléculas gaseosas (p. ej., óxido nítrico, monóxido de carbono)
Endocannabinoides
Glutamato y aspartato
Estos aminoácidos (glutamato y aspartato) son los principales neurotransmisores excitadores del SNC. Están presentes en la corteza, el cerebelo y la médula espinal. En las neuronas, la síntesis de óxido nítrico (NO) aumenta en respuesta al glutamato. El exceso de glutamato puede ser tóxico y aumentar el calcio intracelular, los radicales libres y la actividad de proteinasa. Estos neurotransmisores pueden contribuir a la tolerancia a la terapia con opiáceos y mediar la hiperalgesia.
Los receptores de glutamato (estimulados por el glutamato y menos fuertemente por el aspartato) se clasifican como receptores NMDA (N-metil-d-aspartato) y receptores no NMDA. La fenciclidina (PCP, también conocida como polvo de ángel) y la memantina (utilizada para el tratamiento de la enfermedad de Alzheimer) se unen a receptores de NMDA.
Ácido gamma-aminobutírico
El ácido gamma-aminobutírico (GABA) es el principal neurotrasmisor inhibidor del encéfalo. Es un aminoácido derivado del glutamato, que sufre una descarboxilación por la descarboxilasa de glutamato. Después de la interacción con sus receptores, el GABA es bombeado activamente a las terminaciones nerviosas y es metabolizado. La glicina, parecida al GABA en su acción, está presente sobre todo en las interneuronas (células de Renshaw) de la médula espinal y en los circuitos que relajan los músculos antagonistas.
Los receptores de GABA se clasifican como GABAA (activador de los canales del cloruro) y GABAB (potenciador de la formación de cAMP). Los receptores de tipo GABAA son el punto de acción para diversos fármacos neuroactivos, que incluyen las benzodiazepinas, los barbitúricos, la picrotoxina y el muscimol. El alcohol también se une a los receptores GABA-A. Los receptores de tipo GABA-B son activados por el baclofeno, utilizado para el tratamiento de los espasmos musculares (p. ej., en la esclerosis múltiple).
Serotonina
La serotonina (5-hidroxitriptamina o 5-HT) se sintetiza en los núcleos del rafe y las neuronas de la protuberancia y el tronco encefálico superior. El triptófano es hidroxilado por la triptófano hidroxilasa a 5-hidroxitriptófano y luego descarboxilado a serotonina. Las concentraciones de serotonina están controladas por la captación de triptófano y la monoaminooxidasa (MAO) intraneuronal, que degrada la serotonina. Finalmente, la serotonina es excretada en la orina como el ácido 5-hidroxiindolacético o 5-HIAA.
Los receptores serotoninérgicos (5-HT), de los que hay un mínimo de 15 subtipos, se dividen en 5-HT1 (con 4 subtipos), 5-HT2 y 5-HT3. Sus agonistas selectivos (p. ej., sumatriptán) pueden interrumpir las migrañas. Los inhibidores selectivos de la recaptación de serotonina (ISRS) también se pueden utilizar para tratar varios trastornos de salud mental (p. ej., depresión, ansiedad, trastorno obsesivo compulsivo, trastorno de estrés postraumático).
Acetilcolina
La acetilcolina es el principal neurotransmisor de las neuronas bulboespinales, las fibras preganglionares autónomas, las fibras colinérgicas (parasimpáticas) posganglionares y muchas neuronas del sistema nervioso central (p. ej., ganglios basales, corteza motora). Es sintetizada a partir de la colina y la acetilcoenzima A por la colina acetiltransferasa, y su acción es concluida rápidamente a través de la hidrólisis local a colina y acetato por la acetilcolinesterasa. Las concentraciones de acetilcolina están reguladas por la colina acetiltransferasa y por la captación de colina. Las concentraciones del neurotransmisor están disminuidas en la enfermedad de Alzheimer.
Los receptores colinérgicos se clasifican en nicotínicos N1 (en el músculo esquelético y la unión neuromuscular) o N2 (en los sistemas nerviosos central y periférico, incluidos el parasimpático y el simpático) o muscarínicos M1 a M5 (ampliamente distribuidos en el SNC). Los de tipo M1 se encuentran en el sistema nervioso autónomo, el estriado, la corteza y el hipocampo; los de tipo M2 se encuentran en el sistema nervioso autónomo, el corazón, el músculo liso intestinal, el tronco del encéfalo y el cerebelo.
Dopamina
La dopamina interactúa con los receptores de algunas fibras nerviosas periféricas y de muchas neuronas centrales (p. ej., en la sustancia nigra, el mesencéfalo, el área tegmental ventral y el hipotálamo). El aminoácido tirosina es captado por las neuronas dopaminérgicas y convertido por la tirosina hidroxilasa a 3,4-dihidroxifenilalanina (dopa), que se descarboxila en dopamina por la descarboxilasa de los l-aminoácidos aromáticos. Después de su liberación e interacción con los receptores, la dopamina experimenta un bombeo activo (recaptación) en la terminación nerviosa. La tirosina hidroxilasa y la MAO (que degrada la dopamina) regulan las concentraciones de dopamina en las terminaciones nerviosas.
Los receptores dopaminérgicos se dividen en D1 a D5. Los receptores D3 y D4 desempeñan un papel en el control del pensamiento (limitando los síntomas negativos de la esquizofrenia); la activación de los receptores D2 controla el sistema extrapiramidal. Sin embargo, la afinidad por el receptor no predice la respuesta funcional (actividad intrínseca). Por ejemplo, el ropinirol, que tiene alta afinidad por el receptor D3, presenta una actividad intrínseca a través de la activación de los receptores D2.
Noradrenalina
La noradrenalina es el neurotransmisor de la mayoría de las fibras posganglionares y de muchas neuronas centrales (p. ej., en el locus caeruleus y el hipotálamo). Su precursor, la tirosina, es convertido a dopamina, que es hidroxilada por la dopamina beta-hidroxilasa a noradrenalina. Después de la liberación y la interacción con los receptores, parte de la noradrenalina se degrada mediante la catecol O-metiltransferasa (COMT) y el resto es captado activamente en la terminación nerviosa, donde es degradado por la MAO. La tirosina hidroxilasa, la dopamina beta-hidroxilasa y la MAO regulan las concentraciones intraneuronales de nordrenalina.
Los receptores adrenérgicos son clasificados como alfa-1 (postsinápticos en el sistema simpático), alfa-2 (presinápticos en el sistema simpático y postsinápticos en el encéfalo), beta-1 (en el corazón) o beta-2 (en otras estructuras con inervación simpática).
Endorfinas y encefalinas
Las endorfinas y las encefalinas son opiáceos.
Las endorfinas son polipéptidos grandes que activan muchas neuronas centrales (p. ej., en el hipotalamo, la amígdala, el tálamo y el locus caeruleus). El cuerpo celular contiene un polipéptido grande denominado proopiomelanocortina, precursor de las alfa-endorfinas, las beta-endorfinas y las gamma-endorfinas. La pro-opiomelanocortina es transportada en forma descendente por el axón y escindido en fragmentos; uno es la beta-endorfina, que está presente en las neuronas que se proyectan hacia la sustancia gris periacueductal, las estructuras límbicas y las principales neuronas que contienen catecolaminas en el encéfalo. Después de la liberación y la interacción con los receptores, la beta-endorfina es hidrolizada por las peptidasas.
Las encefalinas incluyen la met-encefalina y la leu-encefalina, que son polipéptidos pequeños presentes en muchas neuronas centrales (p. ej., en el globo pálido, el tálamo, el caudado y la sustancia gris central). Su precursor, la proencefalina, se forma en el cuerpo celular, luego es degradada por peptidasas en los péptidos activos. Estas sustancias se localizan en la médula espinal, donde modulan las señales para el dolor. Los neurotransmisores de las señales para el dolor en el asta posterior de la médula espinal son el glutamato y la sustancia P. Las encefalinas reducen la cantidad de neurotransmisor liberado e hiperpolarizan (vuelven más negativa) la membrana postsináptica, lo que disminuye la generación de potenciales de acción y la percepción del dolor a nivel del giro poscentral. Después de la liberación y la interacción con receptores peptidérgicos, las encefalinas son hidrolizadas en péptidos inactivos más pequeños y aminoácidos. Su inactivación rápida priva a estas sustancias de cualquier utilidad clínica. En su lugar, como analgésicos se emplean moléculas más estables (p. ej., morfina).
Los receptores para endorfina-encefalina (opiáceos) se dividen en mu-1 y mu-2 (que influyen sobre la integración sensitivomotora y la analgesia), delta-1 y delta-2 (que afectan la integración motora, la función cognitiva y la analgesia) y kappa-1, kappa-2 y kappa-3 (que afectan la regulación del equilibrio hidroelectrolítico, la analgesia y el consumo de comida). Los receptores sigma, en la actualidad clasificados fuera de los opiáceos y situados en su mayoría en el hipocampo, se unen a PCP. Los datos más recientes sugieren la presencia de muchos más subtipos de receptores, con sus consecuencias farmacológicas correspondientes. Los componentes del precursor molecular de la proteína receptora pueden reorganizarse durante la síntesis del receptor para producir diversas variantes (p. ej., 27 variantes de corte y empalme del receptor opiáceo mu). Además, 2 receptores pueden combinarse (dimerizarse) para formar un nuevo receptor.
Otros neurotransmisores
Las dinorfinas son un grupo de 7 péptidos con secuencias similares de aminoácidos. Al contrario de las encefalinas, son opiáceos.
La sustancia P, que es un péptido, se encuentra en las neuronas centrales (en la habénula, la sustancia nigra, los ganglios basales, el bulbo raquídeo y el hipotálamo) y presenta una gran concentración en los ganglios de la raíz dorsal. Su liberación es desencadenada por estímulos dolorosos intensos. Modula la respuesta nerviosa al dolor y el estado de ánimo; modula las náuseas y los vómitos a través de la activación de los receptores NK1A que se localizan en el tronco encefálico.
El óxido nítrico (NO) es un gas inestable que interviene en múltiples procesos neuronales. Es generado a partir de la arginina por la NO sintetasa. Los neurotransmisores que aumentan el calcio intracelular (p. ej., sustancia P, glutamato, acetilcolina) estimulan la síntesis de NO en las neuronas que expresan NO sintetasa. El NO puede ser un mensajero intracelular; puede difundir fuera de la célula en una segunda neurona y producir respuestas fisiológicas (p. ej., la potenciación a largo plazo [el reforzamiento de ciertas respuestas presinápticas y postsinápticas, un tipo de aprendizaje]) o favorecer la neurotoxicidad por glutamato (mediada por el receptor de NMDA) (p. ej., en la enfermedad de Parkinson, el accidente cerebrovascular o la enfermedad de Alzheimer). El NO afecta a otros neurotransmisores (p. ej., GABA y acetilcolina) al influir sobre la entrada de calcio en las células para aumentar la liberación de otros neurotransmisores.
Otros neurotransmisores gaseosos adicionales incluyen monóxido de carbono (CO) y sulfuro de hidrógeno (H2S). Estos transmisores se producen en las células de todo el cuerpo (incluido el cerebro). El CO endógeno se genera a partir del metabolismo del hemo y puede participar en procesos que implican la generación de fiebre, la inflamación, la supervivencia celular y el control de la dilatación de los vasos sanguíneos. Varias enzimas están involucradas en la producción de H2S, que se cree que es necesaria para que el cerebro forme y retenga recuerdos.
Las sustancias cuyo rol está peor determinado en la neurotransmisión incluyen las siguientes: histamina, vasopresina, péptido intestinal vasoactivo, carnosina, bradicinina, colecistocinina, bombesina, somatostatina, factor liberador de corticotrofina, neurotensina y posiblemente adenosina.
Los endocannabinoides son neurotransmisores endógenos a base de lípidos que modulan la función cerebral, endocrina y del sistema inmunitario.
Trastornos asociados a defectos de la neurotransmisión
Los trastornos o sustancias que alteran la producción, la liberación, la recepción, la degradación o la recaptación de los neurotransmisores o que modifican el número y la afinidad de los receptores pueden producir síntomas neurológicos o psiquiátricos y causar enfermedad (véase tabla Ejemplos de trastornos asociados a defectos de la neurotransmisión). Los fármacos que modifican la neurotransmisión pueden aliviar muchos de estos trastornos (p. ej., enfermedad de Parkinson, depresión).







