




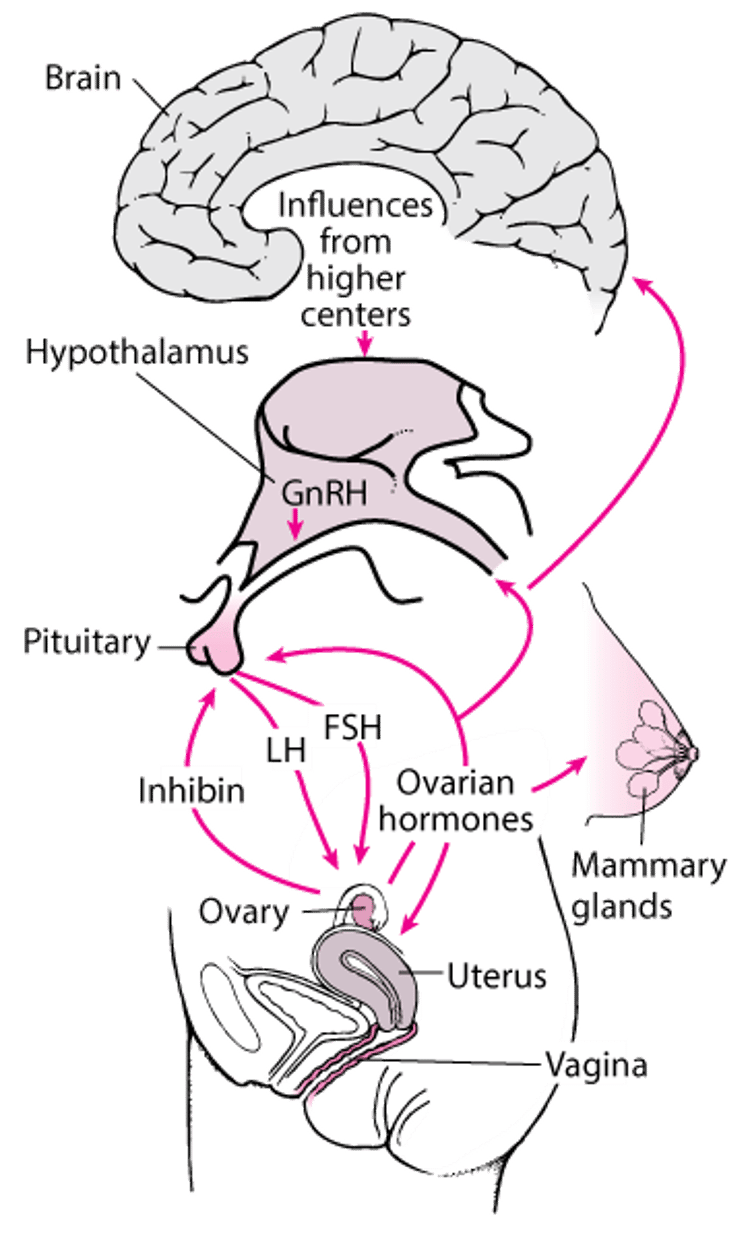
Hormonal interaction between the hypothalamus, anterior pituitary gland, and ovaries regulates the female reproductive system.
The hypothalamus secretes a small peptide, gonadotropin-releasing hormone (GnRH), also known as luteinizing hormone–releasing hormone.
GnRH regulates release of the gonadotropins luteinizing hormone (LH) and follicle-stimulating hormone (FSH) from specialized cells (gonadotropes) in the anterior pituitary gland (see figure The CNS-hypothalamic-pituitary-gonadal target organ axis). These hormones are released in short bursts (pulses) every 1 to 4 hours. LH and FSH promote ovulation and stimulate secretion of the sex hormones estradiol (an estrogen) and progesterone from the ovaries.
Estrogen and progesterone circulate in the bloodstream almost entirely bound to plasma proteins. Only unbound estrogen and progesterone appear to be biologically active. They stimulate the target organs of the reproductive system (eg, uterus, vagina), and the breasts. They usually inhibit (called negative feedback) but, in certain situations (eg, around the time of ovulation), may stimulate gonadotropin secretion.
The central nervous system-hypothalamic-pituitary-gonadal target organ axis
Ovarian hormones have direct and indirect effects on other tissues (eg, bone, skin, muscle). FSH = follicle-stimulating hormone; GnRH = gonadotropin-releasing hormone; LH = luteinizing hormone. |

Puberty
Puberty is the sequence of events in which a child acquires adult physical characteristics and capacity for reproduction. Circulating LH and FSH levels are elevated at birth but fall to low levels within a few months and remain low until puberty. Until puberty, few changes occur in reproductive target organs.
Age of onset of puberty
The age of onset of puberty and the rate of development through different stages are influenced by different factors. Over the last 150 years, the age at which puberty begins has been decreasing, primarily because of improved health and nutrition, but this trend has stabilized. In the US the average age for onset of puberty is 12.5 years, but onset varies by ethnicity (1).
Puberty often occurs earlier than average in moderately obese girls and later than average in severely underweight and undernourished girls (2). Such observations suggest that a critical body weight or amount of fat is necessary for puberty.
Many other factors can influence when puberty begins and how rapidly it progresses. For example, there is some evidence that intrauterine growth restriction, especially when followed by postnatal overfeeding, may contribute to earlier and more rapid development of puberty (3). For example, some evidence suggests that children born small for gestational age have a more advanced progression of puberty (4).
Puberty occurs earlier in girls whose mothers had an earlier age at menarche (5); puberty is also influenced by geographic location and environmental toxicants (6).
Physical changes of puberty
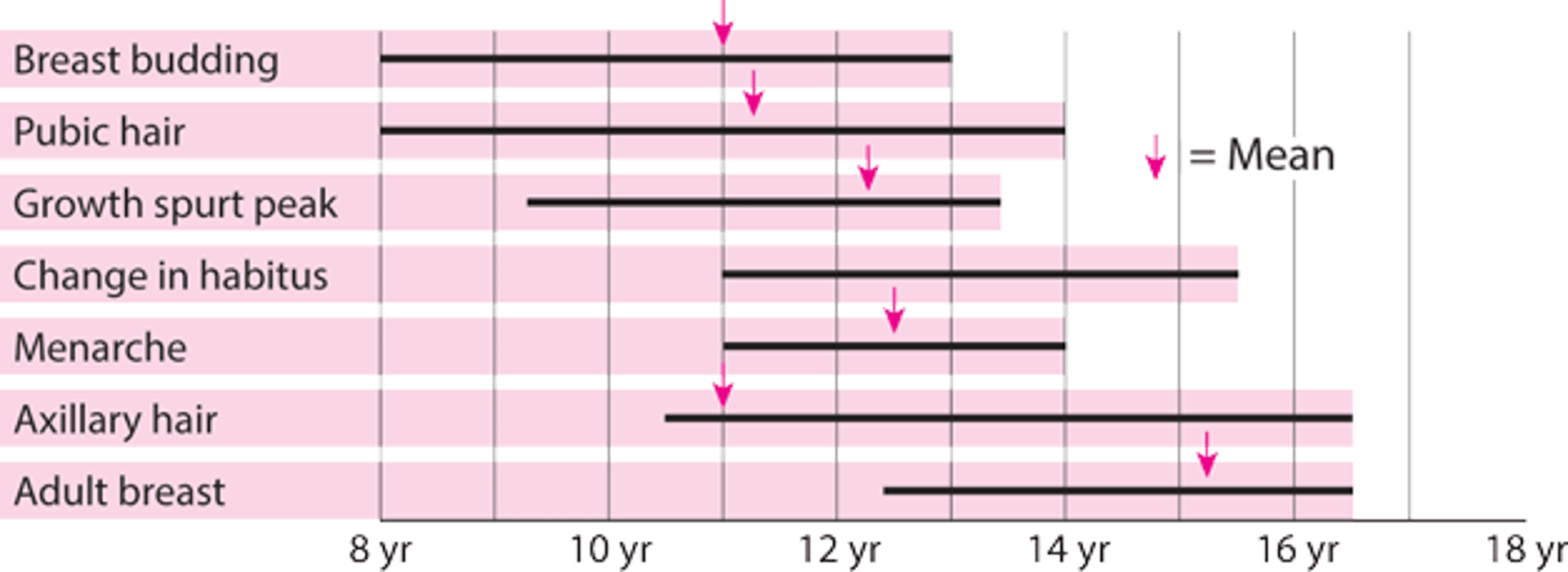
Physical changes of puberty occur sequentially during adolescence (see figure Puberty—when female sexual characteristics develop).
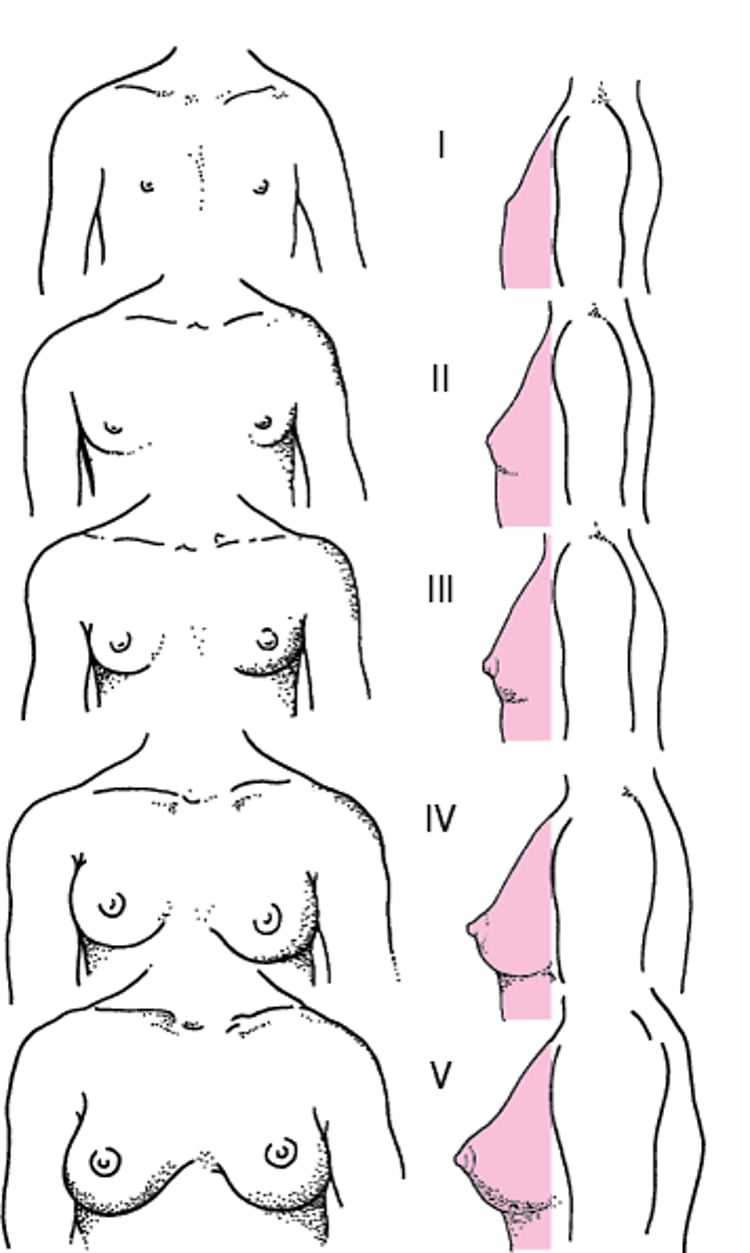
Breast budding (see figure Diagrammatic representation of Tanner stages I to V of breast maturation in girls [7]) and onset of the growth spurt are usually the first changes recognized.
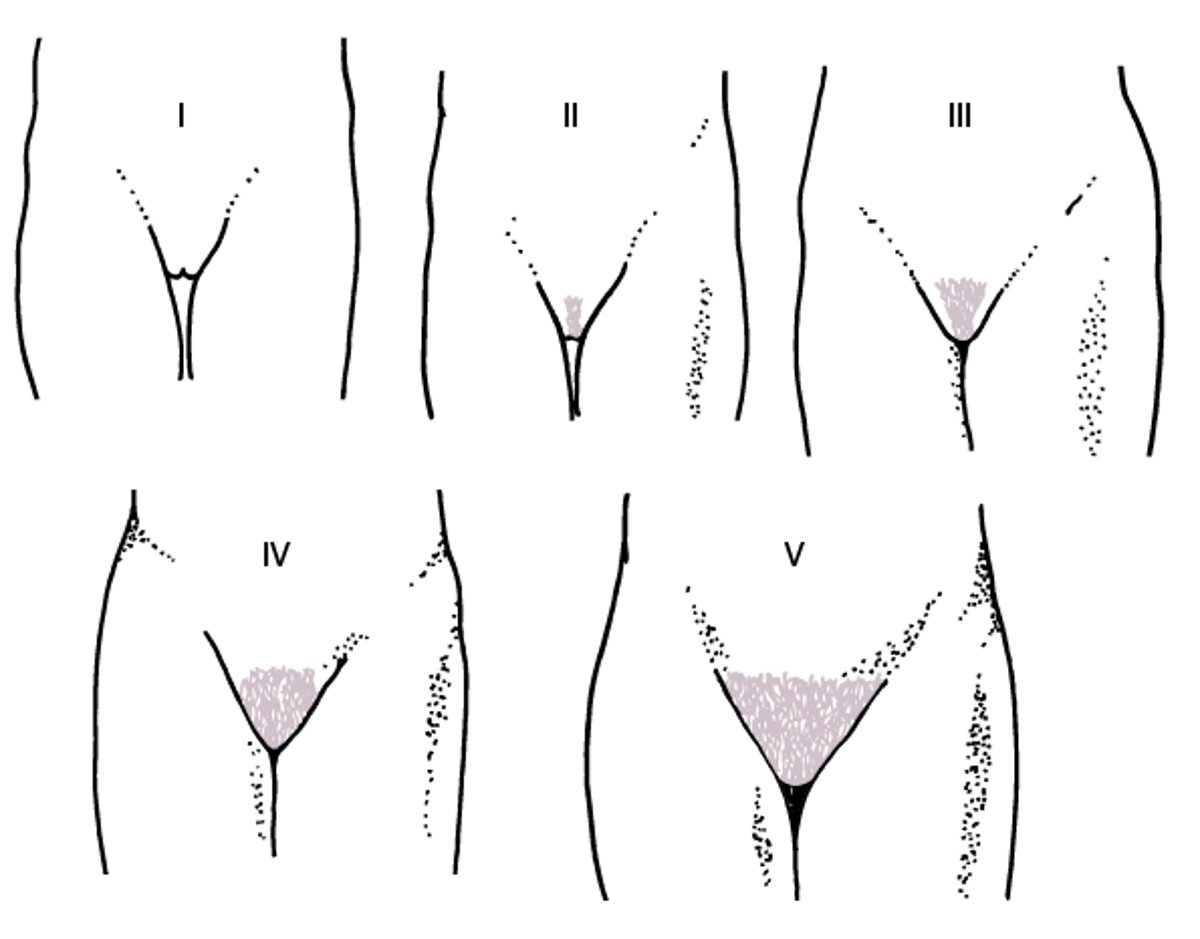
Then, pubic and axillary hair appear (see figure Diagrammatic representation of Tanner stages I to V for development of pubic hair in girls), and the growth spurt peaks.
Menarche (the first menstrual period) occurs about 2 to 3 years after breast budding. Menstrual cycles are usually irregular after menarche and can take up to 5 years to become regular. The growth spurt is limited after menarche. Body habitus changes and the pelvis and hips widen. Body fat increases and accumulates in the hips and thighs.
Mechanisms initiating puberty
Mechanisms initiating puberty are unclear.
Central influences that regulate release of GnRH include neurotransmitters and peptides (eg, gamma-aminobutyric acid [GABA], kisspeptin). Such factors may inhibit release of GnRH during childhood, then initiate its release to induce puberty in early adolescence. Early in puberty, hypothalamic GnRH release becomes less sensitive to inhibition by estrogen and progesterone. The resulting increased release of GnRH promotes LH and FSH secretion, which stimulates production of sex hormones, primarily estrogen. Estrogen stimulates development of secondary sexual characteristics.
Pubic and axillary hair growth is stimulated by the adrenal androgens dehydroepiandrosterone (DHEA) and DHEA sulfate; production of these androgens increases several years before puberty in a process called adrenarche.
Puberty—when female sexual characteristics develop
Bars indicate normal ranges. |

Diagrammatic representation of Tanner stages I to V of breast maturation in girls
From Marshall WA, Tanner JM: Variations in pattern of pubertal changes in girls. Arch Dis Child 44:291–303, 1969; used with permission. |

Diagrammatic representation of Tanner stages I to V for development of pubic hair in girls
From Marshall WA, Tanner JM: Variations in patterns of pubertal changes in girls. Archives of Disease in Childhood 44:291–303, 1969; used with permission. |

Puberty references
1. Anderson SE, Must A: Interpreting the continued decline in the average age at menarche: results from two nationally representative surveys of U.S. girls studied 10 years apart. J Pediatr 147 (6):753–60 2005.doi: 10.1016/j.jpeds.2005.07.016
2. Rosenfield RL, Lipton RB, Drum ML: Thelarche, pubarche, and menarche attainment in children with normal and elevated body mass index. Pediatrics 123 (1):84-8, 2009. doi: 10.1542/peds.2008-0146
3. Darendeliler F: IUGR: Genetic influences, metabolic problems, environmental associations/triggers, current and future management. Best Pract Res Clin Endocrinol Metab 33 (3):101260, 2019. doi: 10.1016/j.beem.2019.01.001 Epub 2019 Jan 22.
4. Veening MA, van Weissenbruch MM, Roord JJ, de Delmemarre-van Waal HA: Pubertal development in children born small for gestational age. J Pediatr Endocrinol Metab 17 (11):1497–505, 2004. doi: 10.1515/jpem.2004.17.11.1497
5. Sørensen S, Brix N, Ernst A, et al: Maternal age at menarche and pubertal development in sons and daughters: A Nationwide Cohort Study. Hum Reprod 33 (11):2043–2050, 2018. doi: 10.1093/humrep/dey287
6. Eckert-Lind C, Busch AS, Petersen JH, et al: Worldwide secular trends in age at pubertal onset assessed by breast development among girls: A systematic review and meta-analysis. JAMA Pediatr 174 (4):e195881, 2020. doi:10.1001/jamapediatrics.2019.5881
7. Marshall WA, Tanner JM: Variations in patterns of pubertal changes in girls. Arch Dis Child 44:291–303, 1969.
Ovarian Follicular Development
A female fetus has a finite number of egg precursors (germ cells). Germ cells begin as primordial oogonia, which migrate from the extraembryonic endoderm of the yolk sac into the undifferentiated genital ridge and become oogonia. Oogonia differentiate into primary oocytes and proliferate markedly by mitosis (single division into two identical diploid cells), resulting in about 7 million oocytes by the 5th month of gestation. During the 3rd month of gestation, some oogonia begin meiosis and arrest in the prophase of meiosis I until puberty. Meiosis occurs in two phases; it results in division into four haploid cells. By the 7th month of gestation, a layer of granulosa cells develops around all viable germ cells and forms a primordial follicle. After the 4th month of gestation, oogonia (and later oocytes) start to be lost spontaneously in a process called atresia; eventually, 99.9% are lost.
Female and male gametogenesis
Gametogenesis is the process of development from primordial germ cells to mature gametes: oogenesis in females and spermatogenesis in males. In both females and males, it begins with diploid germ cells that then undergo mitosis, meiosis, and cytodifferentiation into haploid gametes. |

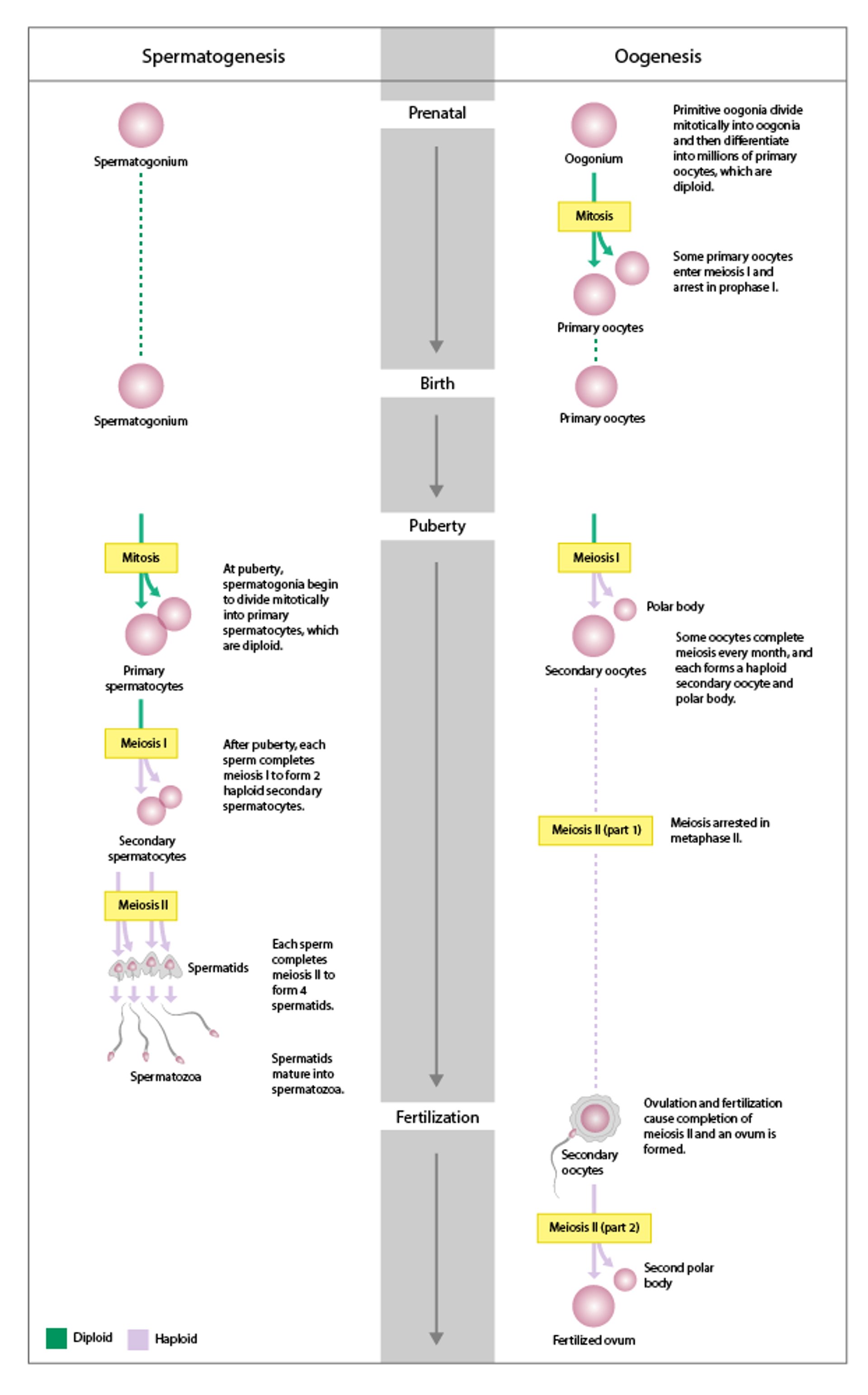
At puberty, the oocyte completes meiosis I to produce one secondary oocyte and one polar body; these cells arrest in metaphase of meiosis II.
FSH induces follicular growth in the ovaries. During each menstrual cycle, 3 to 30 follicles are recruited for accelerated growth. Usually in each cycle, only one follicle achieves ovulation. This dominant follicle releases its oocyte at ovulation and promotes atresia of the other recruited follicles. A single ovum is formed as a result of the two meiotic divisions—one just before ovulation and the second when the sperm penetrates. Polar bodies containing excess genetic material are extruded at each meiotic division. With increasing maternal age, the long time that surviving oocytes spend arrested in meiotic prophase may account for the increased incidence of genetically abnormal pregnancies (1).
Ovarian follicular development reference
1. Jones KT: Meiosis in oocytes: Predisposition to aneuploidy and its increased incidence with age. Hum Reprod Update 14:143–158, 2008.
Menstrual Cycle
Menstruation is the periodic discharge of blood and sloughed endometrium (collectively called menses or menstrual flow) from the uterus through the vagina. It is caused by the rapid decline in ovarian production of progesterone and estrogen that occurs each cycle in the absence of a pregnancy. Menstruation occurs throughout a woman’s reproductive life.
Menopause is defined as 1 year after the last menses.
Normal duration of menses is 4.5 to 8 days (1). Blood loss per cycle averages 30 mL (normal range, 5 to 80 mL) and is usually greatest on the 2nd day. Because patients do not measure menstrual volume, excessively heavy or light menses is determined based on the patient's impression and on the estimated number of pads or tampons used; a saturated pad or tampon absorbs 5 to 15 mL. Menstrual blood does not usually clot (unless bleeding is very heavy), probably because fibrinolysin and other factors inhibit clotting.
The median menstrual cycle length is 28 days (normal range, 24 to 38 days). Generally, variation is maximal and intermenstrual intervals are longest in the years immediately after menarche and immediately before menopause, when ovulation occurs less regularly. In an individual, menses is considered regular when the duration of the shortest and longest cycles vary by ± 2 to 20 days. The duration of the menstrual cycle is counted as the number days from the first day of menses in one cycle to the first day in the next cycle.
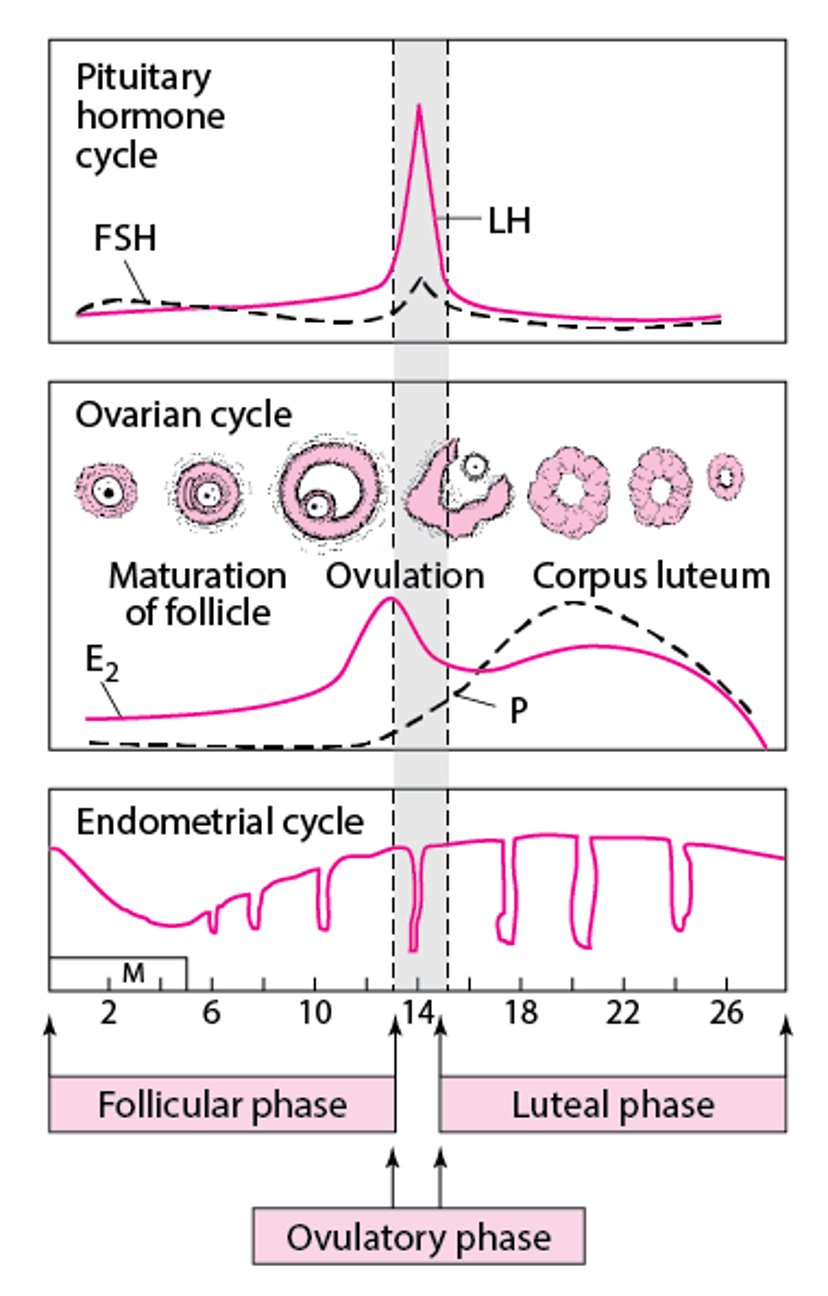
The menstrual cycle can be divided into phases. The ovary proceeds through the following phases:
The endometrium also cycles through phases:
Menstrual
Proliferative
Secretory
Follicular phase
This phase varies in length more than other phases.
In the early follicular phase (first half of the follicular phase), the primary event is
Growth of recruited follicles
At this time, the gonadotropes in the anterior pituitary contain little LH and FSH, and estrogen and progesterone
During the late follicular phaseestrogen, particularly estradiol, increase exponentially.
Ovulatory phase
Ovulation (ovum release) occurs.
Progesterone levels also begin to increase.
progesteroneprogesterone levels continue to increase. The LH surge stimulates enzymes that initiate breakdown of the follicle wall and release of the now mature ovum within about 16 to 32 hours. The LH surge also triggers completion of the first meiotic division of the oocyte within about 36 hours.
Luteal phase
The dominant follicle is transformed into a corpus luteum after releasing the ovum.
The length of this phase is the most constant, averaging 14 days, after which, in the absence of pregnancy, the corpus luteum degenerates.
The corpus luteum secretes primarily progesterone in increasing quantities, peaking at about 25 mg/day 6 to 8 days after ovulation. Progesterone stimulates development of the secretory endometrium, which is necessary for embryonic implantation. Because progesterone is thermogenic, basal body temperature increases by 0.5° C for the duration of this phase.
progesteroneprogesterone levels decrease late in this phase, and the corpus luteum degenerates into the corpus albicans.
Normal Menstrual Cycle

Menstrual cycle reference
1. Fraser IS, Critchley HOD, Broder M, Munro MG: The FIGO recommendations on terminologies and definitions for normal and abnormal uterine bleeding. Semin Reprod Med 29 (5):383–390 2011. doi: 10.1055/s-0031-1287662 Epub 2011 Nov 7.
Cyclic Changes in Other Reproductive Organs
Endometrium
The endometrium, which consists of glands and stroma, has a basal layer, an intermediate spongiosa layer, and a layer of compact epithelial cells that line the uterine cavity. Together, the spongiosa and epithelial layers form the functionalis, a transient layer that is sloughed during menses.
During the menstrual cycle, the endometrium cycles through its own phases:
Menstrual
Proliferative
Secretory
Ovulation occurs at the beginning of the secretory phase of the endometrial cycle. During the ovarian luteal phase, progesteroneprogesterone levels decrease late in the luteal/secretory phase, the stroma becomes edematous, and the endometrium and its blood vessels necrose, leading to bleeding and menstrual flow (menstrual phase of the endometrial cycle). Fibrinolytic activity of the endometrium decreases blood clots in the menstrual blood.
Because histologic changes are specific to the phase of the menstrual cycle, the cycle phase or tissue response to sex hormones can be determined accurately by endometrial biopsy.
Cervix
The cervix acts as a barrier that limits access to the uterine cavity.
During the luteal phase, increasing progesterone levels make the cervical mucus thicker and less elastic, decreasing success of sperm transport.
Menstrual cycle phase can sometimes be identified by microscopic examination of cervical mucus dried on a glass slide; ferning (palm leaf arborization of mucus) indicates increased salts in cervical mucus. Ferning becomes prominent just before ovulation, when estrogen levels are high; it is minimal or absent during the luteal phase. Spinnbarkeit, the stretchability (elasticity) of the mucus, increases as estrogen levels increase (eg, just before ovulation); this change can be used to identify the periovulatory (fertile) phase of the menstrual cycle.
Vagina
During the luteal phase, the number of precornified intermediate cells increases, and the number of leukocytes and amount of cellular debris increase as mature squamous cells are shed.




