



Many factors can affect gene expression (and thus phenotypes). Some cause the expression of traits to deviate from the patterns predicted by Mendelian inheritance.
(See also Overview of Genetics.)
Penetrance and expressivity
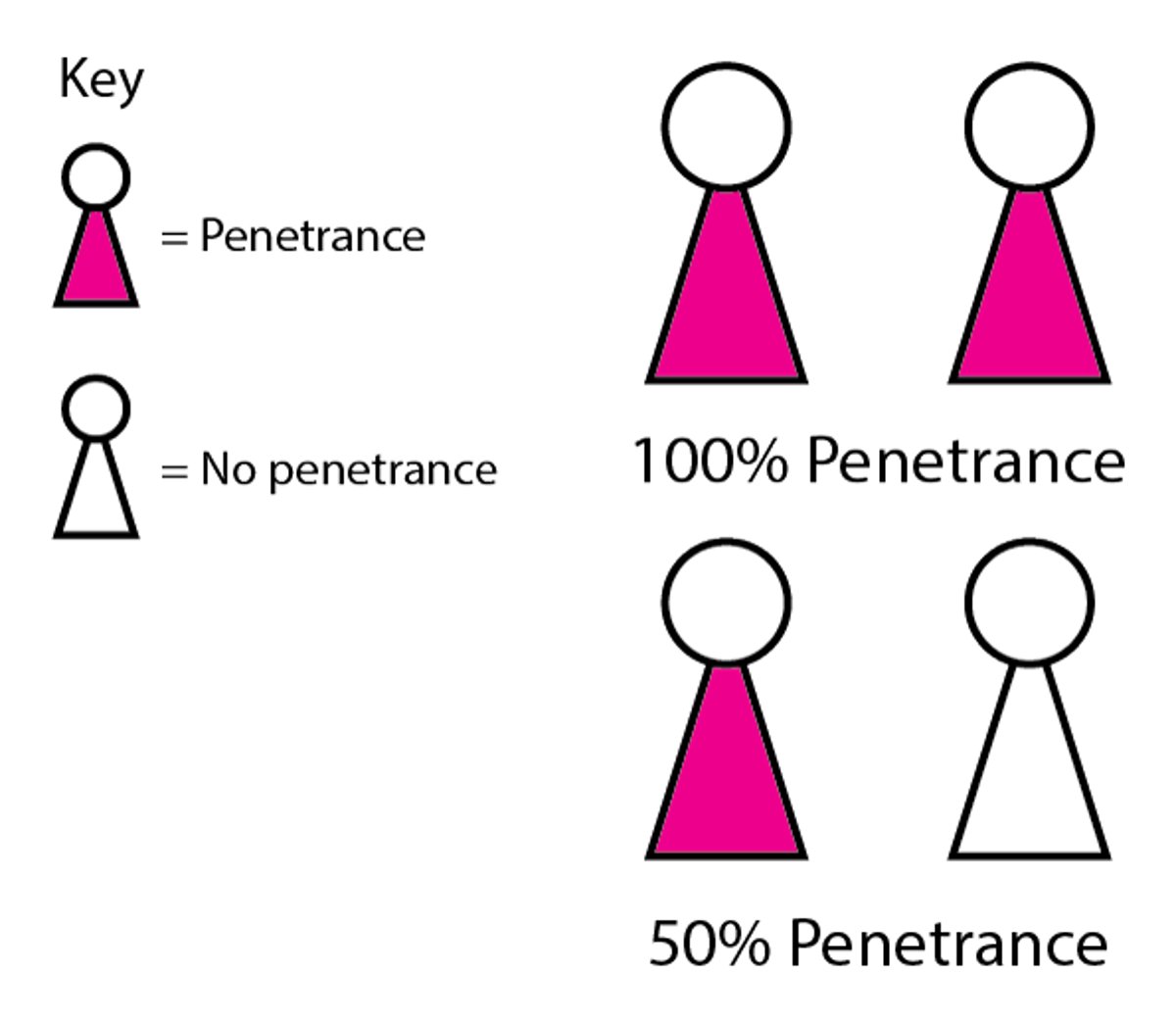
Penetrance is defined as the percentage of people who have the allele and who develop the corresponding phenotype (see figure ). An allele with incomplete (low) penetrance may not be expressed even when the trait is dominant or when it is recessive and the gene responsible for that trait is present on both chromosomes. Penetrance of the same gene may vary from person to person and may depend on a person’s age. Even when an abnormal allele is not expressed (nonpenetrance), the unaffected carrier of the abnormal allele is able to pass it to their children, who may have the clinical abnormality. In such cases, the pedigree appears to skip a generation. However, some cases of apparent nonpenetrance are due to the examiner’s unfamiliarity with or inability to recognize minor manifestations of the disorder. Patients with minimal expression are sometimes considered to have a forme fruste of the disorder.
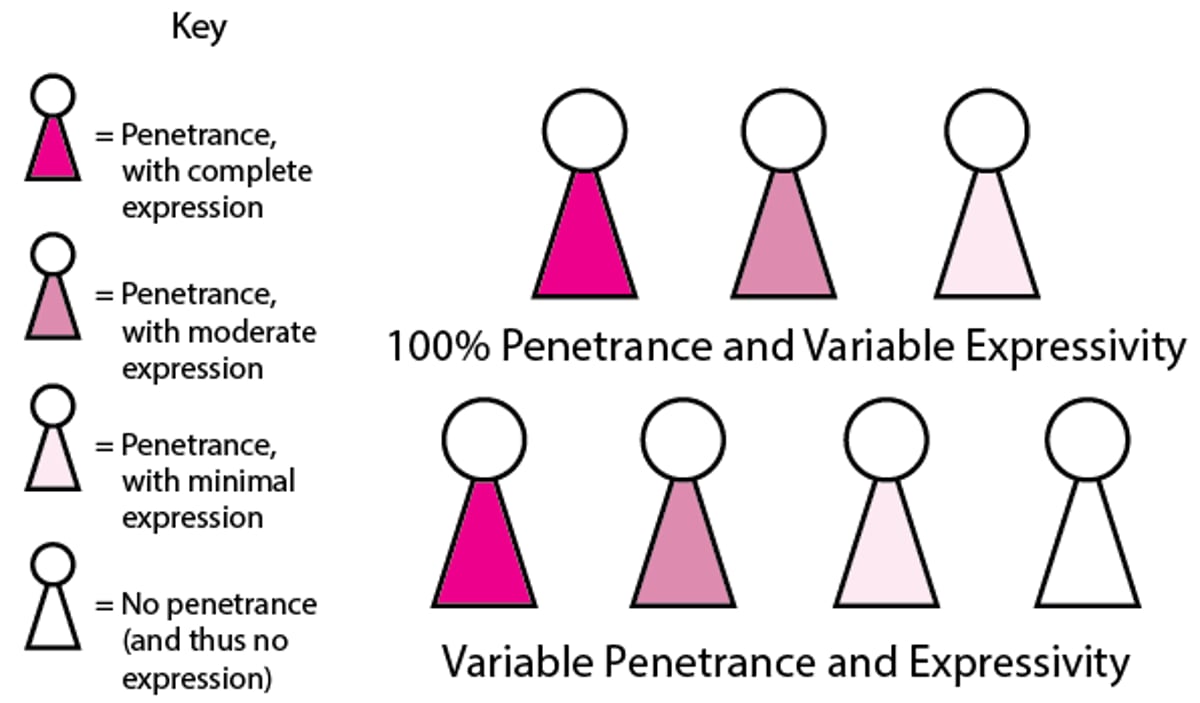
Expressivity is the degree or severity of the phenotype exhibited by individuals who express the trait. It can be quantified as a percentage; eg, when a gene has 50% expressivity, only half the features are present, or the severity is only half of what can occur with full expression. Expressivity may be influenced by the environment and by other genes, so people with the same gene may vary in phenotype. Expressivity can vary even among members of the same family.
Penetrance and Expressivity
How genotype is translated into phenotype depends on penetrance and expressivity. Penetrance refers to whether the gene is expressed or not. That is, it refers to how many people with the gene have the trait associated with the gene. Penetrance may be complete (100%) or incomplete (eg, 50% when only half the people have the trait). Expressivity determines how much the trait affects or how many features of the trait appear in the person. It ranges from complete to minimal, or it may not be present. Various factors, including genetic makeup, exposure to harmful substances, other environmental influences, and age, can affect expressivity. Both penetrance and expressivity can vary: People with the gene may or may not have the trait and, in people with the trait, how the trait is expressed can vary. |


Sex-limited inheritance
A trait that appears in only one sex is called sex-limited. Sex-limited inheritance is distinct from X-linked inheritance, which refers to traits carried on the X chromosome. Sex-limited inheritance, perhaps more correctly called sex-influenced inheritance, refers to special cases in which sex hormones and other physiologic differences between males and females alter the expressivity and penetrance of a gene. For example, premature baldness (known as male-pattern baldness) is an autosomal dominant trait, but such baldness is rarely expressed in females and usually only after menopause.
Genomic imprinting
Genomic imprinting is the differential expression of genetic material depending on whether it has been inherited from the father or mother. For most autosomes, both the paternal and maternal alleles are expressed. However, in < 1% of alleles, expression is possible only from the paternal or maternal allele. For example, expression of the gene for insulin-like growth factor 2 is normally influenced only by the paternal allele.
Genomic imprinting is usually determined by effects that occur normally during the development of gametes. Epigenetic changes such as methylation of DNA or other factors affecting gene expression may cause certain maternal or paternal alleles to be expressed to different degrees. A disorder may appear to skip a generation if genomic imprinting prevents the causative allele from being expressed. Defective imprinting, such as abnormal activation or silencing of alleles, can result in clinical disorders (eg, Prader-Willi syndrome, Angelman syndrome).
Codominance
Codominant alleles are both observed. Thus, the phenotypic expression of heterozygotes is distinct from that of either homozygote. For example, if a person has 1 allele coding for blood type A and 1 allele coding for blood type B, the person expresses both blood types (blood type AB).
Chromosomal inactivation
(See also Lyon hypothesis [X inactivation].)
In females, who have 2 (or, with sex chromosomal abnormalities, > 2) X chromosomes (except in eggs), all but one of the X chromosomes is inactivated; ie, most of the alleles on that chromosome are not expressed. Which chromosome is inactivated is determined randomly, individually in each cell and early in fetal life; such a phenomenon is called lyonization. Sometimes, it is the X from the mother that is inactivated, and sometimes it is the X from the father. Sometimes, most of the X chromosome inactivation comes from one parent—called skewed X inactivation. Either way, once inactivation has taken place in a cell, all descendants of that cell have the same X inactivation.
However, some alleles on the inactive X chromosome can be expressed. Many of these alleles are on chromosomal regions corresponding to regions of the Y chromosomes (and are thus called pseudoautosomal regions because both males and females receive 2 copies of these regions).
Key Points
If a trait in a pedigree appears to skip a generation, consider incomplete penetrance, incomplete expression, and (less likely) genomic imprinting.
Phenotypes can also be modified by sex-limited inheritance, genomic imprinting, codominance of alleles, and X chromosome inactivation.





