



Hemostasis, the arrest of bleeding from an injured blood vessel, requires the combined activity of:
Vascular factors
Platelets
Plasma coagulation factors
Regulatory mechanisms counterbalance the tendency of clots to form. Hemostatic abnormalities can lead to excessive bleeding or thrombosis.
Vascular Factors of Hemostasis
Vascular factors reduce blood loss due to trauma through local vasoconstriction (an immediate reaction to injury) and compression of injured vessels by extravasation of blood into surrounding tissues. Vessel wall injury triggers the attachment and activation of platelets and the generation of fibrin polymers from fibrinogen; platelets and fibrin combine to form a clot.
Platelets
Various mechanisms, including endothelial cell nitric oxide and prostacyclin, promote blood fluidity by preventing platelet aggregation and dilating intact blood vessels. These mediators are no longer produced when the vascular endothelium is disrupted. Under these conditions, platelets adhere to the damaged intima and form aggregates. Initial platelet adhesion is due to long strings of von Willebrand factor (VWF) that have been previously secreted by endothelial cells and anchored to exposed subendothelial collagen at the sites of endothelial damage. VWF binds to receptors on the platelet surface membrane (glycoprotein Ib/IX). Platelets anchored to the vessel wall undergo activation. During activation, platelets release mediators of aggregation, including adenosine diphosphate (ADP) from storage granules.
Other biochemical changes resulting from activation include:
Hydrolysis of membrane phospholipids
Inhibition of adenylate cyclase
Mobilization of intracellular calcium
Phosphorylation of intracellular proteins
Arachidonic acid is converted to thromboxane A2; this reaction requires platelet cyclooxygenase and is inhibited irreversibly by aspirin and reversibly by many NSAIDs (nonsteroidal anti-inflammatory drugs).
Adenosine diphosphate, thromboxane A2, and other mediators induce activation and aggregation of additional platelets on the injured endothelium. Platelet receptors for ADP include the P2Y12 receptor, which sends signals to suppress adenylate cyclase, decreases cyclic adenosine monophosphate (cAMP) levels, and promotes activation of the glycoprotein IIb/IIIa receptor (assembled on the activated platelet surface membrane from glycoproteins IIb and IIIa). Fibrinogen binds to the glycoprotein IIb/IIIa complexes of adjacent platelets, connecting them into aggregates.
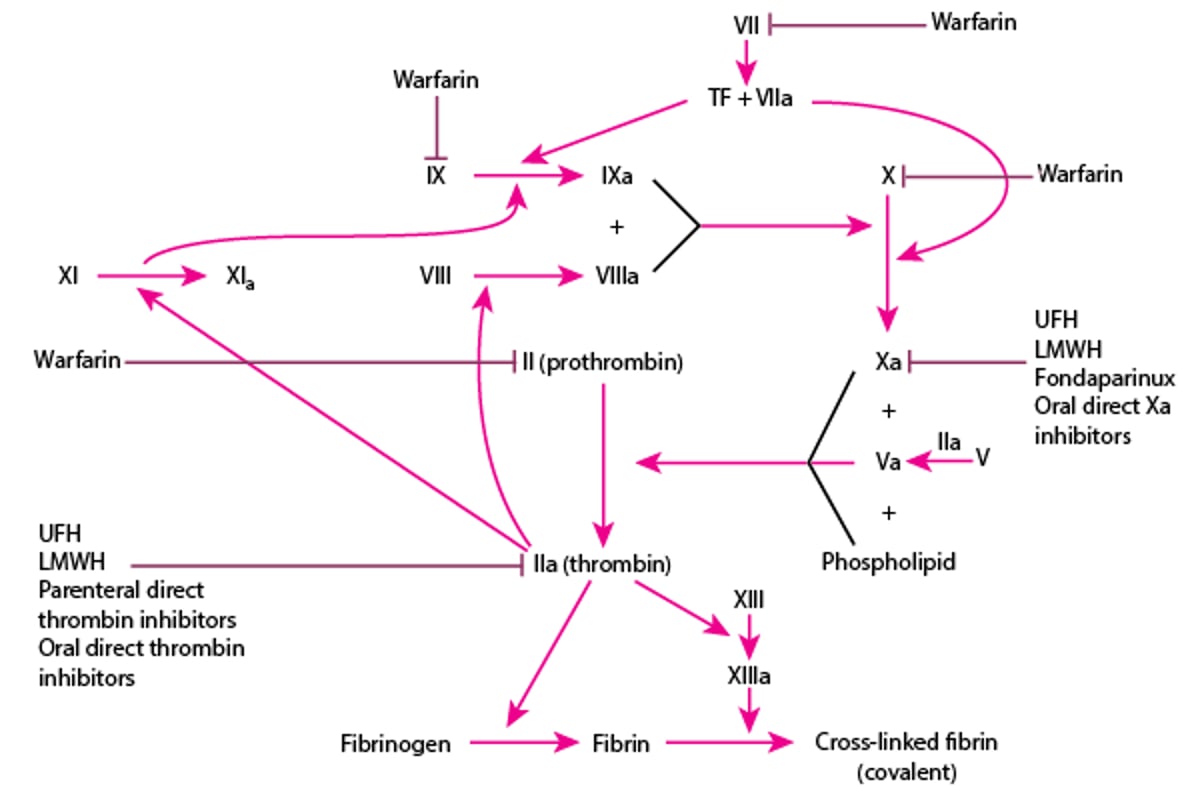
Platelets provide surfaces for the assembly and activation of coagulation complexes and the generation of thrombin. Thrombin converts fibrinogen into fibrin monomers, and the fibrin monomers polymerize into fibrin polymers that bind aggregated platelets into platelet-fibrin hemostatic plugs. Thrombin also activates factor VIII and factor V, key cofactors in the tenase (Factors IXa and VIIIa) and prothrombinase (Factors Xa and factor Va) complexes, respectively. Thrombin also feeds back and activates factor XI to amplify activation of the intrinsic pathway leading to greater factor X activation and ultimately greater fibrin formation. Thrombin also activates factor XIII which catalyzes covalent crosslinks between fibrin polymers, strengthening the fibrin clot (see figure ).
Plasma Coagulation Factors
Coagulation factors interact on platelet and endothelial cell surfaces to produce thrombin, which converts fibrinogen to fibrin. By radiating from and anchoring the hemostatic plug, fibrin strengthens the clot.
In the intrinsic pathway, factor XII, high molecular weight kininogen, prekallikrein, and activated factor XI (factor XIa) interact to produce factor IXa from factor IX. Factor IXa then combines with factor VIIIa and procoagulant phospholipid (present on the surface of activated platelets, endothelial cells, and tissue cells) to form a complex that activates factor X.
In the extrinsic pathway, factor VIIa and tissue factor (TF) directly activate factor X (and perhaps also factor IX—see figure and table ).
The coagulation factors are produced in the liver with the exception of factor VIII, which is synthesized in liver sinusoidal cells and endothelial cells outside the liver. Tissue factor expression is normally restricted to perivascular cells so that extrinsic pathway activation only occurs in the event of vessel wall injury.
Pathways in Blood Coagulation


Components of Blood Coagulation Reactions
Factor Number or Name | Synonym | Purpose |
|---|---|---|
Plasma factors | ||
I | Fibrinogen | A precursor of fibrin monomers (and polymers) |
II | Prothrombin | The precursor of thrombin Thrombin converts fibrinogen to fibrin; activates soluble factors V, VIII, XI, and XIII; and binds to thrombomodulin to activate protein C Is vitamin K–dependent |
V | Proaccelerin | Is activated to factor Va, a cofactor for the enzyme factor Xa in the factor Xa/factor Va/phospholipid complex, which cleaves prothrombin to thrombin Is present in alpha granules in platelets Factor Va inactivated by activated protein C in complex with free protein S |
VII | Proconvertin | Binds to tissue factor and is then activated to form the enzymatic component of the factor VIIa/tissue factor complex, which activates factor X and, possibly, factor IX Is vitamin K–dependent |
VIII | Antihemophilic globulin | Is activated to factor VIIIa, a cofactor for the enzyme factor IXa in the factor IXa/factor VIIIa/phospholipid complex, which activates factor X Is a large cofactor protein (as is factor V) Is secreted bound to von Willebrand factor multimers from the Weibel-Palade bodies of endothelial cells (factor VIII also circulates with von Willebrand factor) As factor VIIIa, is inactivated by activated protein C in complex with free protein S (as is factor Va) |
IX | Christmas factor | Is activated to factor IXa, the enzyme of the factor IXa/factor VIIIa/phospholipid complex, which activates factor X Is vitamin K–dependent |
X | Stuart-Prower factor | Is activated to factor Xa, the enzyme of the factor Xa/factor Va/phospholipid complex, which cleaves prothrombin to thrombin Is vitamin K–dependent |
XI | Plasma thromboplastin antecedent | Is activated to factor XIa, which activates factor IX |
Prekallikrein | Fletcher factor | Participates in a reciprocal reaction in which it is activated to kallikrein by factor XIIa Kallikrein, catalyzes further activation of factor XII to factor XIIa Circulates as a biomolecular complex with high molecular weight kininogen |
High molecular weight kininogen | Fitzgerald factor | Circulates as a bimolecular complex with prekallikrein and facilitates prekallikrein activation to kallikrein |
XII | Hageman factor | When activated to factor XIIa by surface contact, kallikrein, or other factors, it activates prekallikrein and factor XI, triggering the intrinsic coagulation pathway in vitro |
XIII | Fibrin stabilizing factor | When activated by thrombin, catalyzes formation of peptide bonds between adjacent fibrin monomers to strengthen and stabilize fibrin clots |
Protein C | — | Is activated by thrombin bound to surface membrane-bound thrombomodulin (CD141); then proteolyzes and inhibits (in the presence of free protein S and phospholipid) the cofactor activity of factor VIIIa and factor Va Is vitamin K–dependent |
Protein S | — | Circulates in plasma as free protein S and as protein S bound to C4b-binding protein of the complement system Functions in its free form as a cofactor for activated protein C Is vitamin K–dependent |
Cell surface factors | ||
Tissue factor | Tissue thromboplastin | Is a protein that is present in the membranes of certain tissue cells, including perivascular fibroblasts, endothelial cells, boundary epithelial cells (eg, epithelial cells of the skin, amnion, and gastrointestinal and genitourinary tracts), glial cells of the nervous system, monocytes, macrophages, and some tumor cells It is exposed to flowing blood during injury or inflammation, binds factor VIIa, and initiates the extrinsic coagulation pathway |
Procoagulant phospholipid | — | Acidic phospholipid (primarily phosphatidyl serine) present on the surface of activated platelets, endothelial cells, and other tissue cells Is a component of the factor IXa/VIIIa/phospholipid complex which activates factor X and of the factor Xa/factor Va/phospholipid complex which activates prothrombin |
Thrombomodulin | CD141 | Is an endothelial cell surface binding site for thrombin which, when bound to thrombomodulin, activates protein C |
Activation of the intrinsic or extrinsic pathway activates the common pathway, resulting in formation of the fibrin clot. Three steps are involved in common pathway activation:
Prothrombinase is generated on the surface of activated platelets, endothelial cells, and tissue cells. Prothrombinase is a complex of an enzyme, factor Xa, and a cofactor, factor Va, on a procoagulant phospholipid surface.
Prothrombinase cleaves prothrombin to thrombin.
Thrombin induces the generation of fibrin monomers and polymers from fibrinogen and activates soluble factors V, VIII, and XI. Thrombin also activates factor XIII, an enzyme that catalyzes formation of stronger, covalent bonds between adjacent fibrin monomers.
Calcium ions are required in most thrombin-generating reactions and, therefore, calcium-chelating agents (eg, citrate, ethylenediaminetetraacetic acid) are used in vitro as anticoagulants. Vitamin K–dependent clotting factors (factors II, VII, IX, and X) normally bind to phospholipid surfaces through calcium bridges to function in blood coagulation. Coagulation reactions cannot occur properly in the absence of vitamin K. Vitamin K-dependent coagulation regulatory proteins include protein C and protein S.
Although the coagulation pathways are helpful in understanding mechanisms and laboratory evaluation of coagulation disorders, in vivo coagulation does not require factor XII, prekallikrein, or high molecular weight kininogen. Patients with hereditary deficiencies of these factors have no bleeding abnormality. Patients with hereditary factor XI deficiency may have a mild to moderate bleeding disorder. In vitro, soluble factor XI can be activated by thrombin. There is, however, no consistent relationship between plasma factor XI levels and the likelihood or extent of bleeding. Soluble factor IX can be activated in vitro both by factor XIa and factor VIIa/tissue factor complexes.
In vivo, initiation of the extrinsic pathway occurs when injury to blood vessels brings blood into contact with tissue factor on membranes of cells within and around the vessel walls. This contact with tissue factor generates factor VIIa/tissue factor complexes that activate factor X (and possibly factor IX). Factor IXa, combined with its cofactor, factor VIIIa, on phospholipid membrane surfaces also generates factor Xa. Factor X activation by factor IXa/VIIIa complexes is required for normal hemostasis. This requirement for factors VIII and IX explains why hemophilia type A (deficiency of factor VIII) or type B (deficiency of factor IX) results in bleeding. Factor X activation by factor VIIa/tissue factor complexes in the extrinsic coagulation pathway does not generate sufficient thrombin (and fibrin) to prevent bleeding in patients with severe hemophilia A or B.
Regulation of Coagulation
Several inhibitory mechanisms prevent activated coagulation reactions from amplifying uncontrollably, causing extensive local thrombosis or disseminated intravascular coagulation. These mechanisms include:
Inactivation of coagulation factors
Fibrinolysis
Hepatic clearance of activated clotting factors
Inactivation of coagulation factors
Plasma protease inhibitors (antithrombin, tissue factor pathway inhibitor [TFPI], alpha 2-macroglobulin, and heparin cofactor II) inactivate coagulation enzymes. Antithrombin inhibits thrombin, factor Xa, factor XIa, and factor IXa. Tissue factor pathway inhibitor inhibits factor Xa as well as the tissue factor/factor VIIa (TF/FVIIa) complex.
Two vitamin K–dependent proteins, protein C and free protein S, form a complex that inactivates factors VIIIa and Va by proteolysis. Thrombin, when bound to a receptor on endothelial cells (thrombomodulin [CD141]), activates protein C. Endothelial protein C receptor binds protein C and facilitates its activation by the thrombin-thrombomodulin complex. Activated protein C, in combination with free protein S proteolyzes and inactivates factors VIIIa and Va.
In addition to normally present inactivators, there are a number of anticoagulant medications that potentiate the inactivation of coagulation factors (see figure ).
Heparin enhances antithrombin activity and triggers the release of TFPI. Unfractionated heparin (UFH) and low molecular weight heparins (LMWHs) enhance activity of antithrombin to inactivate factors IIa (thrombin) and Xa. They also stimulate release of TFPI from the endothelial cells which inhibits factor Xa and the TF/FVIIa complex. LMWHs include enoxaparin, dalteparin, and tinzaparin.
Warfarin is a vitamin K antagonist. It inhibits regeneration of the active form of vitamin K and, therefore, inhibits generation of functional forms of the vitamin K–dependent clotting factors II, VII, IX, and X (as well as proteins C and S).
Fondaparinux, a small, synthetic molecule containing the essential pentasaccharide portion of the heparin structure, enhances antithrombin inactivation of factor Xa but not factor IIa (thrombin).
Parenteral direct thrombin inhibitors include argatroban and bivalirudin.
The direct oral anticoagulants include the thrombin inhibitor (dabigatran) and the factor Xa inhibitors (apixaban, rivaroxaban, edoxaban). For a more detailed discussion on the use of these medications, including risks, benefits, and reversal agents, see atrial fibrillation, deep venous thrombosis (DVT), and pulmonary embolism (PE).
Anticoagulants and Their Sites of Action
LMWH = low molecular weight heparin; TF = tissue factor; UFH = unfractionated heparin. |

Fibrinolysis
Fibrin deposition and lysis are normally balanced to maintain temporarily, and subsequently remove, the hemostatic seal during repair of an injured vessel wall. The fibrinolytic system dissolves fibrin by means of plasmin, a proteolytic enzyme. Fibrinolysis is activated by plasminogen activators such as tissue plasminogen activator (TPA) which are released from vascular endothelial cells in response to thrombin and shear stress. Plasminogen activators and plasminogen (from plasma) bind to fibrin, and plasminogen activators cleave plasminogen into plasmin (see figure ). Plasmin then proteolyzes fibrin into soluble fibrin degradation products that are swept away in the circulation and metabolized by the liver.
Fibrinolytic Pathway
Fibrin deposition and fibrinolysis must be balanced during repair of an injured blood vessel wall. Injured vascular endothelial cells release plasminogen activators (tissue plasminogen activator, urokinase), activating fibrinolysis. Plasminogen activators cleave plasminogen into plasmin, which dissolves clots. Fibrinolysis is controlled by plasminogen activator inhibitors (PAIs; eg, PAI-1) and plasmin inhibitors (eg, alpha 2-antiplasmin). |

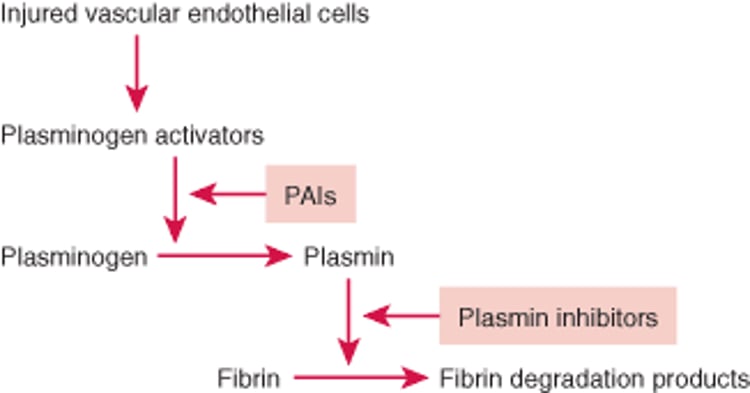
There are several plasminogen activators:
Tissue plasminogen activator (TPA), from endothelial cells, is a poor activator of plasminogen when free in solution but an efficient activator when bound to fibrin in proximity to plasminogen. TPA is released from endothelial cells in response to thrombin or shear stress.
Urokinase exists in single-chain and double-chain forms with different functional properties. Single-chain urokinase cannot activate free plasminogen but, like TPA, can readily activate plasminogen bound to fibrin. A trace concentration of plasmin cleaves single-chain to form double-chain urokinase, which activates plasminogen in solution, as well as plasminogen bound to fibrin. Epithelial cells that line excretory passages (eg, renal tubules, mammary ducts) release urokinase, which is the physiologic activator of fibrinolysis in these channels.
Streptokinase, a bacterial product not normally found in the body, is another potent plasminogen activator.
Streptokinase, urokinase, and recombinant TPA (alteplase) have all been used therapeutically to induce fibrinolysis in patients with acute thrombosis.
Regulation of fibrinolysis
Fibrinolysis is regulated by plasminogen activator inhibitors (PAIs) and plasmin inhibitors that slow fibrinolysis. PAI-1, the most important PAI, inactivates TPA, and urokinase and is released from vascular endothelial cells and activated platelets. The primary plasmin inhibitor is alpha 2-antiplasmin, which quickly inactivates any free plasmin escaping from clots. Some alpha 2-antiplasmin is also cross-linked to fibrin polymers by the action of factor XIIIa during clotting. This cross-linking may prevent excessive plasmin activity within clots.
Tissue plasminogen activator and urokinase are rapidly cleared by the liver, which is another mechanism of preventing excessive fibrinolysis.
Drug Information for the Topic





